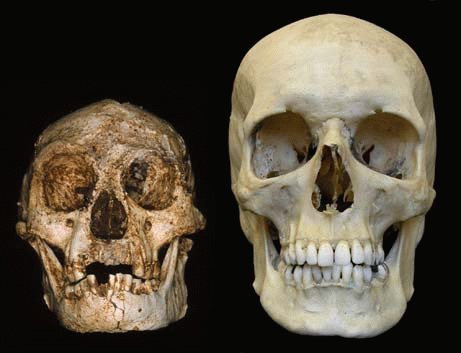
 |
| Comparaison entre le crâne d'Homo floresiensis (à gauche) et celui d'Homo sapiens (à droite) |
 |
| Homo floresiensis (vue d'artiste) |
Le mystère de l'homme de Flores
Un documentaire de Annamaria Talas, Simon Nasht (52')(Australie, 2008)
Un documentaire de Annamaria Talas, Simon Nasht (52')(Australie, 2008)
________________________________________________________
Une nouvelle théorie alternative de l'évolution humaine...
(En cours d'élaboration. Dernière mise à jour : 19 octobre 2012)
Je souhaite présenter sur ce blog une proposition alternative quant à l'évolution, depuis le Miocène, de la lignée ayant donné naissance à l'Humanité. Rien que cela!
N'ayant aucune qualification particulière sanctionnée par un diplôme dans les domaines concernés (paléoanthropologie, génétique, etc.), ce que je reconnais bien volontiers, je ne cherche à convaincre personne, mais tout au plus, en tant qu'amateur s'appuyant sur un certain nombre de données récentes et de travaux reconnus ou plus controversés, à intriguer et à faire s'interroger d'éventuels lecteurs quant à la plausibilité de l'hypothèse présentée. Chacun reste libre de n'y voir qu'une plaisante fiction... Et peut-être effectivement n'est-ce là qu'une fiction... ou pas.
Alors commençons par évoquer en quelques lignes le paradigme qui a dominé la réflexion des scientifiques, pendant une vingtaine d'années et qui, malgré son abandon progressif depuis le début du XXIe siècle, risque fort d'imprégner encore longtemps l'esprit de Monsieur-Tout-le-Monde en étant véhiculé de façon dominante par les représentations populaires et le système éducatif.
Une thèse abandonnée... ou presque
 |
| Une "image d'Epinal" naïve et... totalement fausse |
Le problème est que, au cours des dernières décennies, se sont multipliées les découvertes d'hominidés plus ou moins anciens montrant que l'évolution de la lignée humaine depuis des âges très reculés se présente non pas de façon linéaire, mais buissonnante ; tout comme celle des autres espèces animales d'ailleurs, l'Homme ne faisant pas exception en la matière. Le schéma d'évolution allant du DAC (dernier ancêtre commun entre les grands singes anthropoïdes africains et l'espèce humaine) à l'Homo sapiens, en passant par les australopithèques, les Homo habilis, puis les Homo erectus ne tient plus, et cela les spécialistes le reconnaissent aujourd'hui bien volontiers. Les tentatives d'intégrer à ce schéma les hominidés découverts plus ou moins récemment sont vouées à l'échec, alors que, par exemple, on sait aujourd'hui avec certitude que vers 1,5 ou 2 millions d'années BP, en Afrique de l'Est et du Sud, cohabitaient diverses sous-espèces d'australopithèques, des paranthropes, des Homo habilis, des Homo rudolfensis et des Homo ergaster. Rien ne prouve d'ailleurs qu'une espèce dont on n'a trouvé pour l'instant que peu de restes fossiles, datés de 3,5 à 3,2 millions d'années, Kenyanthropus platyops, n'a pas survécu jusque-là.
On retrouve ce schéma d'une évolution buissonnante même à une date beaucoup plus récente dans le cas de la lignée Homo, puisque, c'est là aussi aujourd'hui une certitude, au moins quatre humanités différentes coexistaient encore sur la planète voilà seulement 40 000 ou 50 000 ans : l'Homme de Néandertal, l'Homme de Denisova, l'Homme de Flores (mais nous verrons que, selon mon hypothèse, celui-ci n'appartient sans doute pas stricto sensu au genre Homo) et Homo sapiens. Peut-être même cinq, voire six, si on y ajoute Homo soloensis, découvert en 1931 sur l'île de Java, dont la datation reste encore très controversée (135 000 ans? 50 000 ans? 25 000 ans?) ou les énigmatiques Hommes dits "du Cerf rouge" dont les fossiles, datés de 14 000 à 11 000 ans BP ont été découverts en Chine entre 1979 et 1989,mais n'ont commencé à être étudiés qu'en 2008.
D'autre part, comment expliquer, dans le cadre du paradigme exposé ci-dessus, que l'on retrouve des formes anciennes (Ardipithecus, Orrorin tugenensis, Toumaï) autant, sinon plus bipèdes que des plus récentes (les australopithèques)? Avec en plus, dans le cas de Toumaï (Sahelanthropus tchadensis), daté de 7 millions d'années, une incohérence manifeste par rapport à la date estimée de séparation entre la lignée humaine et celle des chimpanzés (5 à 6 millions d'années BP, selon les travaux en biologie moléculaire). Incohérence aussi, dans le cas de Homo floresiensis qui se serait éteint vers -18 000 ans à peine. Ce sidérant hominidé avait la capacité crânienne d'un australopithèque, à peine supérieure à celle d'un chimpanzé. Il avait aussi des pieds assez similaires à ceux des Homo habilis ayant vécu 2 millions d'années auparavant, voire à ceux des australopithèques, avec une longueur évoquant celle des grands singes. Et pourtant, ainsi que le montre l'ensemble de sa morphologie osseuse, notamment la position du trou occipital, il était bipède et par ailleurs, était, semble-t-il, à l'origine d'une industrie lithique (voir ici, pour un bon résumé des polémiques concernant ce fossile qui déboussole les chercheurs).
Presque en même temps que s'effondrait ce schéma linéaire de l'évolution humaine, une théorie largement dominante pendant deux décennies était elle aussi remise en cause par les faits : "l'East Side Story". Cette théorie supposait que le passage de témoin des primates préhumains quadripèdes arboricoles à la lignée humaine de plus en plus bipède s'était effectuée à partir de 8 millions d'années BP, du côté est du Grand Rift Africain lorsqu'une grande faille s'est formée du nord au sud dans cette région de l'Afrique (la Rift Valley), séparant d'un côté, à l'est, des paysages évoluant vers la savane avec régression des espaces boisés et de l'autre, à l'ouest, une persistance de la forêt. Or, la découverte, en 1995, d'Abel (Australopithecus bahrelghazali) dans un lieu situé aujourd'hui au Tchad, à l'ouest de la Rift Valley, puis en 2011, au Tchad également, de Toumaï, un hominidé encore plus ancien, invalide définitivement la théorie, comme Yves Coppens lui-même l'a désormais reconnu.
Diverses hypothèses alternatives
La situation actuelle dans le domaine de la paléoanthropologie est bien loin des affirmations assurées des vingt ou trente dernières décennies. La mise sur la touche des australopithèques comme ancêtres directs de l'Homme fait que l'on en revient plus ou moins, presque à contrecœur, à des interrogations que l'on croyait dépassées depuis les années 1960 (voir la présentation des thèses de l'époque dans l'ouvrage Homogenesis, de Claude Stoll, 2011, pp. 276 et sq.) sur l'hominisation et notamment sur l'ancienneté et les modalités de l'acquisition de la bipédie humaine. Au point qu'un dossier spécial du périodique Philosophie Magazine (n°35, décembre 2009 / janvier 2010) clamait voilà un an de cela, par le biais d'un gros titre particulièrement racoleur, comme si l'on avait trouvé le Saint-Graal : "Le singe descend de l'homme!". Titre racoleur, donc, mais aussi et surtout totalement erroné et particulièrement susceptible d'introduire de nouvelles croyances trompeuses dans les esprits des non-initiés. En effet, le singe ne descend aucunement de l'homme, pas plus que l'homme ne descend du singe. L'"homme" et le "singe" (en tout cas les grands singes anthropoïdes) descendent en fait d'un ancêtre commun primate. Mais, contrairement à ce que la pensée majoritaire soutenait jusque-là, il semble se confirmer que ce primate encore inconnu était bien plutôt bipède que quadrupède et que notre espèce Homo est probablement plus proche morphologiquement parlant, par certains aspects, de ce primate ancien que les grands anthropoïdes actuels (bien que nous soyons une espèce spécialisée dans la marche et dans la course, donc adepte d'une bipédie quasi exclusive, ce que n'était certainement pas l'ancêtre primate qui devait certainement être, lui, peu spécialisé). Même les pontes représentants de la pensée jusque-là majoritaire, comme Yves Coppens (présent dans le magazine cité ci-dessus) ou Pascal Picq paraissent progressivement venir à cette idée, laquelle, il faut le souligner, n'est pourtant en rien une nouveauté, puisque Yvette Deloison, chercheuse au CNRS et ancienne élève d'Yves Coppens, avait eu le courage de la soutenir, contre vents et marées, dès la fin des années 1990, notamment dans son ouvrage remarquable publié en 2004 (Préhistoire du piéton. Essai sur les nouvelles origines de l'homme, Plon).
 |
| Yvette Deloison, CNRS |
La thèse présentée ici, orientée dans ce sens d'une grande ancienneté de la bipédie dans la lignée ayant abouti à l'espèce humaine, s'inspire principalement d'éléments de réflexion tirés de trois modèles théoriques plus ou moins controversés, tout en prétendant apporter une approche inédite et originale sur le sujet :
- Le premier modèle, diffusé sous le nom de "théorie de la bipédie initiale", a été développé par le zoologue spécialisé en ichtylogie, François de Sarre, à partir des conceptions de l'anatomiste Max Westenhoffer et du zoologue Serge Frechkop, reprises par le zoologue (et père de la cryptozoologie), Bernard Heuvelmans. Sans souscrire complètement, loin de là, à cette théorie, notamment dans certains de ses développements que j'estime totalement fantaisistes (je reviendrai sur cela plus loin), la découverte fortuite de cette hypothèse, voilà deux ou trois ans sur Internet, a titillé ma curiosité et m'a poussé à chercher plus loin que ce que l'on m'avait toujours enseigné sur le sujet.
- Le second modèle porte le nom de "théorie du singe (ou du primate) aquatique" (en anglais : Aquatic Ape Theory). Elle a été proposée à l'origine par le zoologue Sir Alistair Hardy, et a ensuite été développée et popularisée principalement par la journaliste anglaise Elaine Morgan. On en trouvera une présentation par E. Morgan elle-même (sous-titres en français) en suivant le lien :
 |
| Elaine Morgan |
- Le troisième est celui auquel mon hypothèse emprunte le plus. Il s'agit de la théorie de la "bipédie originelle", proposée par Yvette Deloison, déjà citée.
Pour présenter en quelques mots la théorie de la "bipédie initiale" développée par François de Sarre, celle-ci, prenant un complet contre-pied par rapport à la perspective dominante, soutient qu'un organisme marin de l'ère primaire, structuré verticalement par une chorde dorsale, serait devenu, en s'adaptant à la vie terrestre sous une forme initialement bipède, l'ancêtre de tous les vertébrés. Soit. L'hypothèse, présentée comme cela apparaît intéressante et pas moins plausible que les thèses officielles accréditant l'idée d'un organisme marin développé sur un plan horizontal sortant de l'eau en rampant, tel que je peux l'estimer en tout cas avec mes maigres connaissances de base en biologie, surtout en anatomie, en embryologie (puisque la théorie de F. de Sarre se base surtout sur ces deux disciplines) et en évolution du monde vivant. Et la réponse par avance de l'auteur aux éventuelles critiques concernant l'absence de traces fossiles, jusqu'à présent, de cet organisme très ancien me semblent plutôt justifiées (voir ici pour un résumé de la théorie de la bipédie initiale par son auteur, F. de Sarre).
Par contre, le problème est que François de Sarre n'en reste pas là : il soutient que cet animal, qu'il baptise du nom d'"homoncule marin archéopage" aurait eu une forme quasi-humaine et que, par voie de conséquence, ce serait une espèce humaine ou "para-humaine" très ancienne, héritière en ligne directe de cet organisme, qui aurait donné naissance à toutes les autres espèces de vertébrés.
 |
| L'homocule marin archéopage selon F.de Sarre |
 |
| Arbre phylogénétique selon le modèle de la Bipédie initiale, selon F. de Sarre |
Il avance même que ces "Hommes" auraient bâti, non seulement au cours du Quaternaire, mais aussi au Tertiaire et au Secondaire, de brillantes civilisations qui se seraient effondrées de façon cyclique, en conséquence peut-être de cataclysmes planétaires, et que les survivants auraient évolué par "déshumanisation" vers diverses formes animales... Outre le fait que ce serait là un bon scénario de roman de science-fiction, cette théorie me paraît comporter un vice logique rédhibitoire : du fait de la loi de l'irréversibilité de l'évolution (loi de Dollo), le pied humain, hautement spécialisé dans la marche et dans la course, ne peut avoir donné naissance au pied des grands singes quadrumanes, même en passant par le stade australopithèque et encore moins aux pattes des diverses familles de vertébrés. Ce problème logique est analogue, en inversé, à celui rencontré dans l'hypothèse classique : la main morphologiquement "primitive" de l'espèce humaine et son pied spécialisé dans la bipédie permanente ne peuvent dériver de ceux d'un ancêtre quadrupède arboricole, éventuellement brachiateur, dont les membres auraient été adaptés à ce mode de locomotion.
A ce titre, l'hypothèse proposée par la chercheuse Yvette Deloison sous le nom de "théorie de la bipédie originelle", semble, elle, beaucoup plus cohérente, en avançant l'idée d'un "protohominoïde bipède" non-spécialisé apparu vers 15 millions d'années BP comme origine de la lignée commune au genre humain et aux grands singes anthropoïdes, et c'est cette théorie qui est véritablement le point de départ de la thèse soutenue ici, bien que, selon moi, ce "protohominoïde bipède" a en réalité une ancienneté bien supérieure, puisque je place son apparition à environ 30 millions d'années BP.
 |
| Illustration de l'hypothèse du protohominoïde bipède selon Y. Deloison Schéma extrait de la page ci-dessous |
Mais il est temps, maintenant, de présenter mon hypothèse.
Le plus lointain ancêtre de la lignée humaine?
Selon moi, tout commence, contrairement aux théories classiques, à l'Eocène, vers 40 millions d'années BP, en Asie du sud-est, avec l'apparition des premiers singes anthropoïdes à partir de la souche représentée par les fossiles de primates haplorhiniens très primitifs, datés de 45 millions d'années BP (famille des eosimiidae proche de celles des omomyidés qui a donné les tarsiers actuels) trouvés en Chine en 1994 (voir le dossier présenté par Jean-Jacques Jaeger, chercheur du CNRS, qui abonde en ce sens : http://www.cnrs.fr/cw/dossiers/Origine/carte/anthropoides.htm). Le nouveau taxon, colonisant les forêts tropicales humides, se répand rapidement dans le sud de l'Asie sous la forme d'un buisson d'espèces (Ganlea, Bahinia, Pondaungia, Siamopithécus et Amphipithecus, trouvés au Myanmar - Birmanie - et en Thaïlande). Un de ces primates, Afrasia djijidae, trouvé en 2005 en Birmanie, mérite une attention particulière : en effet, il possède un proche "cousin" africain, Afrotarsius libycus, découvert en Libye, approximativement du même âge (37 millions d'années BP pour le premier, 38 ou 39 millions d'années BP pour le second) un peu plus évolué que lui (voir ici). Cela montre que le taxon s'est répandu très tôt d'Asie en Afrique où l'on retrouve, dès -35 millions d'années, un foisonnement de nouvelles espèces (oligopithecus, aegyptopithecus et propliopithecus). C'est à cette époque qu'un groupe de cette famille de primates anthropoïdes se retrouve, de façon assez énigmatique (on pense à une dérive de plusieurs jours sur des radeaux naturels flottants à partir des côtes africaines), en Amérique du sud (encore isolée à l'époque du futur continent nord-américain) où ils évolueront de façon séparée pour donner la branche des platyrhiniens (représentée aujourd'hui par les singes du Nouveau Monde), alors que les primates haplorhiniens d'Afrique et d'Eurasie donnaient naissance, eux, à la branche des catarhiniens.
Toutefois, contrairement là encore aux théories habituelles, c'est encore, selon moi, en Asie du sud-est que se produit l'étape suivante, probablement à l'Oligocène, entre 25 et 30 millions d'années BP, avec une mutation entraînant la perte de la queue. La nouvelle espèce qui se développe alors a probablement des membres supérieurs approximativement de la même longueur que les inférieurs, ressemblant grosso modo, on peut le supposer, à un gibbon, mais avec des bras beaucoup plus courts que ceux d'aujourd'hui*. Les individus de cette nouvelle espèce passent certainement une partie de leur existence à terre à pratiquer la bipédie (mais une bipédie différente de la nôtre, avec des pieds sans voûte plantaire) en milieu semi-aquatique (marais, mangroves, forêts tropicales inondées) qu'ils colonisent préférentiellement, tout en adoptant la quadripédie sur terrain sec, notamment en cas de fuite, et en se réfugiant au besoin dans les arbres, sans pour autant y pratiquer la brachiation. C'est le "protohominoïde bipède" supputé par Yvette Deloison comme étant à l'origine des Hominoïdés. Il colonise à son tour les zones tropicales d'Eurasie méridionale et d'Afrique.
[* Edit du 19.06.2012 : Ayant cherché quels singes actuels ont le plus d'affinités avec l'eau, je me suis aperçu qu'une espèce asiatique est tout particulièrement connue pour cela : le nasique (genre nasalis, famille des cercopithécidés), vivant à Bornéo. C'est également un des rares primates (avec aussi, entre autres le gibbon) à utiliser de façon assez régulière la bipédie. Voir à ce propos la page wikipédia dont voici un extrait :
"Le nasique est le seul non-humain, avec le gibbon et le grand pangolin, à utiliser la station debout pour ses déplacements. Des nombreux groupes ont été filmés marchant sur leur pattes arrières en file indienne longeant des sentiers forestiers, les femelles transportant leurs petits sur leurs hanches. Le nasique est un nageur hors-pair. Il nage d'île en île et certains ont même été pris dans un filet de bateau de pêche à plus d'1,5 kilomètre de la côte."
 |
| Nasique mâle (nasalis larvatus - source wikipédia) |
Est-il possible que le primate à l'origine, par mutation, du "protohominoïde" bipède ait été proche, voilà 25 ou 30 millions d'années de l'ancêtre du nasique - et aussi d'un cousin du nasique, beaucoup plus rare actuellement (en voie de disparition, même), classé auparavant dans le genre Nasalis et aujourd'hui dans le genre Simias (Simias concolor, autrefois appelé Nasalis concolor, connu aussi sous le nom vernaculaire de Simakobu), dotée d'une courte queue glabre, qui survit aujourd'hui dans quatre îles indonésiennes, proches de Sumatra, de l'archipel Mentawaï? Simias concolor (qui mesure de 50 à 60 cms et dont les quatre membres sont de même longueur, comme le signale ce site consacré aux primates, qui en donne la description complète en anglais) constituerait alors l'évolution préférentiellement arboricole des descendants non mutants de cet ancêtre commun, alors que son cousin, sans doute proche d'apparence mais sans queue et préférentiellement bipède, avec des compétences en milieu aquatique similaires à celles du nasique actuel, évoluait lui à terre (dans les mangroves, les forêts humides, en milieu marécageux et sur les bords de rivière), vers la forme du gibbon actuel (mais avec des bras courts) pour donner naissance à la lignée des hominoïdes. Fin de l'Edit].
 |
| Simakobu (Simias concolor) |
 |
| Simakobu (Simias concolor). La petite queue glabre de ce singe est visible sous la branche sur laquelle ce spécimen est assis |
| Simakobu (Simias Concolor) |

[Edit du 22.06.2012 : En poursuivant mes recherches, je viens de m'apercevoir que le genre Nasalis est le seul parmi les primates autres que les grands singes anthropoïdes a posséder 48 chromosomes dans son génome... Coïncidence... Ou pas?]
[Edit du 25.06.2012 : Après quelques jours de réflexion étayée par des recherches sur Internet, je suis désormais convaincu que mon intuition exposée ci-dessus est la bonne, et je pense même avoir trouvé le "berceau" du protohominoïde bipède, qui serait d'ailleurs, selon moi, le même que celui de son successeur, ancêtre direct d'Homo floresiensis : l'île de Siberut, la plus grande et la plus septentrionale des quatre îles habitées de l'archipel Mentawaï, à l'ouest de Sumatra.
 |
| Carte de l'archipel mentawaï, appartenant à l'ensemble géographique "sundaland" en Indonésie |
 |
| Carte de Siberut |
Siberut est en effet une île très particulière, du point de vue de la faune et de la flore, car on y observe une extraordinaire biodiversité, avec de nombreuses espèces endémiques inconnues ailleurs, notamment pour ce qui concerne les quatre espèces de primates qui y survivent tant bien que mal aujourd'hui, menacées d'extinction : une espèce de macaque (Macaca siberu), un semnopithèque (Presbytis potenziani siberu), un gibbon (Hylobates klossii), ainsi que l'espèce cousine du nasique dont il a été question plus haut (Simias concolor). Certaines espèces sont considérées comme représentantes de la faune "primitive" de la région géographique à laquelle appartient l'archipel Mentawaï (à savoir la plaque "sunda" ou "sundaland" dont il sera question plus loin), alors que d'autres ont évolué vers des formes bien distinctes de celles présentes sur le continent. Pour ce qui concerne les îles Mentawaï en général et Siberut en particulier, certains chercheurs ont proposé l'hypothèse que ces îles ont joué un rôle crucial dans l'évolution des écosystèmes en Asie du sud-est. Brandon Jones, par exemple, a suggéré que les primates de l'archipel Mentawaï - préservés lors des périodes glaciaires, car ces îles ont toujours conservé un climat et une biotope tropical, tout au long de l'histoire mouvementée de cette région du monde - auraient pu contribuer, en période interglaciaire, au repeuplement de Sumatra dont la faune aurait été, elle, décimée par le froid, notamment pour ce qui concerne les primates (Brandon-Jones 1998).
Si cette hypothèse est controversée, il est certain que, alors qu'en période glaciaire, le niveau de la mer étant beaucoup plus bas qu'aujourd'hui, les îles de la sonde constituaient les sommets d'une sorte de péninsule reliée au continent asiatique, tourmentée par une intense activité volcanique : le sundaland, l'archipel Mentawaï, apparu par soulèvement tectonique au cours du tertiaire, n'a connu, lui que de rares moments de rattachement, lorsque le niveau marin était au plus bas, grâce à un pont terrestre reliant Siberut à Sumatra (la dernière fois, entre 1 million et 500 000 ans BP - les deux îles étant distantes aujourd'hui d'une centaine de kilomètres). Siberut et ses îles soeurs ont donc connu de très longues périodes d'isolement total, pendant lesquelles une faune et une flore originale ont pu se développer... Pour cette raison, Siberut apparaît comme l'endroit idéal pour l'apparition du "protohominoïde bipède" mais aussi de l'ancêtre direct d'Homo floresiensis. D'autant plus que l'environnement de cette île constitue, et ce depuis des millions d'années, un milieu particulièrement propice à la naissance de ces nouvelles espèces, car, conformément à mon hypothèse au sujet de ce berceau de la lignée des hominoïdes, une grande partie de la surface de Siberut est recouverte de forêts tropicales primaires humides, avec d'importantes zones marécageuses et de très vastes espaces de mangrove sur la côte est...
Voilà donc, selon moi, ce qui a dû se passer entre 30 et 20 millions d'années BP dans cette région du monde (voir plus bas pour des hypothèses plus détaillées) :
- 30 millions d'années BP : Alors que la famille des cercopithécidés est en voie de constitution, un primate de la sous-famille des colobinés elle aussi en émergence, connaît une évolution originale sur le plateau continental Sunda alors isolé du reste du continent : sans doute morphologiquement proche de l'une et de l'autre des espèces actuelles de Nasique, cumulant cependant une longue queue et des aptitudes bipèdes et aquatiques comparables à celles du nasique actuel (Nasalis larvatus), mais avec un appendice nasal peu développé (comme Simias concolor).
- Lors d'une période glaciaire lors de laquelle Siberut est reliée à Sumatra, dans un intervalle de temps compris entre 30 et 25 millions d'années BP, ce primate s'installe dans l'archipel Mentawaï. Puis l'île est à nouveau isolée pour plusieurs centaines de milliers (plusieurs millions?) d'années pendant lesquelles le primate en question voit sa queue se réduire nettement en taille, selon une évolution en deux branches différentes : une espèce préférentiellement arboricole qui conserve une petite queue quasiment glabre (Simias concolor), et une espèce complètement dépourvue de queue, préférentiellement bipède et semi-aquatique : le "protohominoïde bipède" cher à Yvette Deloison.
- 25 millions d'années BP, un nouveau pont terrestre permet le passage de cette nouvelle espèce vers Sunda, puis vers le continent asiatique. Ensuite, Siberut est à nouveau isolée et le protohominoïde bipède y continue son évolution originale qui va le mener vers 20 millions d'années BP à donner naissance à l'ancêtre direct d'Homo floresiensis.
- 20 millions d'années BP : Cet ancêtre peut à son tour rejoindre et coloniser le plateau continental Sunda, puis le continent asiatique... Fin de l'Edit].
 |
| Les gibbons actuels adoptent spontanément la marche bipède lors de leurs (rares) descentes au sol |
Même des cercopithèques actuels autres que le nasique peuvent adopter occasionnellement la bipédie en milieu
semi-aquatique (voir l'exemple ci-dessus, chez des babouins filmés lors d'une traversée de rivière)
Dans cette vision des choses, c'est donc au début du Miocène, vers 20 millions d'années BP, que se produit une nouvelle mutation, majeure pour l'apparition future de la lignée humaine : elle touche avant tout les fonctions cognitives et langagières, très probablement du fait d'une ou de plusieurs modifications sur le gène FOXP2. Toutefois, il ne s'agit sans doute pas encore de la version humaine de ce gène. En conséquence, le cerveau et la boîte crânienne des individus de cette nouvelle espèce (dont le génome compte probablement 48 chromosomes) augmentent de taille, mais surtout le nombre de neurones se multiplient ainsi que les connexions synaptiques, tandis que se développent des lobes frontaux et temporaux impliqués dans la cognition, l'activité intellectuelle, la mémoire, la planification et le langage.
A quoi ressemble alors l'individu de cette nouvelle espèce, que l'on pourrait - pourquoi pas? - désigner, par commodité pour la suite de cet exposé, du nom de Primo ancestor? Selon moi, il est possible d'en avoir une idée car des fossiles morphologiquement proches ont bel et bien été trouvés. Même si les vestiges en questions ne sont pas ceux d'individus de la souche originelle d'il y a 20 millions d'années mais sont beaucoup plus récents, je suis convaincu que, grâce à eux, on peut aujourd'hui avoir une idée assez précise de ce à quoi Primo ancestor devait ressembler. Car je propose l'hypothèse qu'Homo floresiensis - puisqu'il s'agit de lui - est un descendant de cette souche originelle, dont des petits groupes, n'ayant presque pas évolué au cours de ces vingt derniers millions d'années, auraient survécu jusqu'au paléolithique (et peut-être même jusqu'à l'époque contemporaine) dans certains coins reculés de l'Asie du sud-est, berceau de son espèce.
Si l'on prend Homo floresiensis comme modèle, Primo ancestor aurait donc mesuré environ un mètre de haut pour un poids de 16 à 28 kgs. Sa capacité crânienne aurait été de 300 à 400 cm3. Il aurait été faiblement prognathe, avec une dentition très semblable à celle des représentants du genre Homo qui apparaîtront des millions d'années plus tard (notamment de petites canines) et un bourrelet sus-orbitaire réduit et nettement séparé en deux parties. La morphologie de ses membres supérieurs lui auraient permis de grimper facilement aux arbres sans pour autant être brachiateur. Ses pieds, très longs relativement au tibia et au fémur, proches en cela de ceux des grands singes actuels, étaient dépourvus de voûte plantaire (http://www.hominides.com/html/actualites/homo-floresiensis-pied-femur-bipede-0194.php).
Par ailleurs, si l'on accepte l'hypothèse que l'Homme de Florès est aussi l'Ebu Gogo (trad. : "la Grand-Mère qui mange n'importe quoi") décrit dans les légendes encore vivaces aujourd'hui au sein des populations humaines locales, voici ce que l'on peut savoir de lui, et donc, selon moi, de Primo ancestor : velu, il mesurait effectivement environ un mètre de haut, avait un ventre proéminent, des oreilles décollées et de longs cheveux. Les femelles avaient, semble-t-il, de longs seins pendant. Les légendes confirment que, malgré sa démarche bipède maladroite, il était capable de grimper aux arbres avec une grande agilité, et surtout, il avait, semble-t-il, un appétit féroce (des Ebu Gogo auraient même enlevé et dévoré des bébés humains dans les temps historiques selon les villageois colportant ces récits, ce qui aurait nécessité leur extermination... Mais c'est peut-être là simplement une explication a posteriori pour justifier cette éradication par Homo sapiens). Selon ces légendes, bien qu'il ait disposé d'un langage, avec par ailleurs la capacité de répéter les phrases prononcées par les Homo sapiens de façon écholalique, il n'aurait pas su se servir d'outils... Pourtant,on a découvert une industrie lithique associée aux fossiles d'Homo florensiensis et ailleurs dans la région, qui daterait pour les plus anciennes pièces de 840 000 ans, sans que l'on puisse les attribuer à un hominidé local, hormis à l'Homme de Flores. L'explication de l'"énigme" est pour moi, la suivante : sous la pression d'Homo Sapiens, présent dans l'île à partir de 11 000 ans BP, Homo florensiensis a probablement commencé une régression évolutive, avec peut-être d'une part une diminution de la bipédie au profit de l'arboricolisme, mais surtout un abandon progressif de certaines technologies culturelles maîtrisées jusque-là, comme l'industrie lithique rudimentaire.
 |
| La "dame de Florès" (vue d'artiste) |
Pour l'instant, continuons de dérouler notre scénario qui n'est peut-être qu'une belle histoire... ou peut-être pas
Primo ancestor va rapidement devenir dominant au sol par rapport à ses congénères non-mutants en développant ses capacités cognitives, organisationnelles et sociales, et va progressivement reléguer ces derniers dans les arbres où ils vont se spécialiser pour donner naissance à la lignée conduisant aux gibbons, houloks et siamangs actuels, brachiateurs aux très longs membres supérieurs. Au fil des millions d'années, la nouvelle espèce va, elle, se répandre vers l'ouest au cours du Miocène, en s'adaptant à des environnements divers et en égrenant localement des espèces-filles composées d'individus plus moins bipèdes ou plus ou moins arboricoles. En Asie, ce seront les lufengpithèques, khoratpithèques, ramapithèques et sivapithèques, représentants d'une lignée se séparant aux alentours de 15 ou 16 millions d'années BP en deux branches. La première donnera le gigantopithèque aujourd'hui disparu (à moins qu'il n'en subsiste quelques petits groupes dans les montagnes himalayennes, où il serait alors connu localement sous le nom de Dzu-Teh, autrement dit le grand Yéti. Un descendant du gigantopithèque pourrait être aussi le fameux "Bigfoot" d'Amérique du Nord, à moins que ce dernier ne soit issu d'un autre descendant de cette branche : le très robuste meganthrope - meganthropus paleojavanicus - dont on a retrouvé quelques restes osseux, essentiellement des maxillaires, dans les années 1940-1950 à Sangiran sur l'île de Java).
La seconde branche aboutira à l'orang-outan actuel, mais aussi à l'espèce d'orang-outan préférentiellement bipède (Pongo hooijeri) dont des restes fossiles ont été découverts au Vietnam et ailleurs en Asie du sud-est et qui pourrait bien vivre encore dans certaines régions reculées d'Asie (ce serait en ce cas, par exemple, le Meh-Teh - le petit Yéti - de l'Himalaya, mais il pourrait bien aussi vivre encore aujourd'hui à Sumatra ou il serait confondu avec l'Orang pendek dont il sera question plus loin).
En Europe, les descendants de Primo ancestor seront notamment des griphopithèques, des ankarapithèques, des ouranopithèques, des graecopithèques, et des oréopithèques. Mais les plus intéressants parmi ces "singes du Miocène" européens descendants plus ou moins arboricoles de Primo ancestor sont certainement Pierolapithecus catalaunicus, daté de 13 millions d'années BP, et Anoiapithecus brevirostris, surnommé Lluc (12 millions d'années BP), découverts tous deux en Espagne, qui semblent l'aboutissement occidental du périple européen de Primo ancestor, avant le passage de celui-ci en Afrique, aux alentours de 8 millions d'années BP, pour donner naissance à la lignée des grands singes africains, dont l'homme.
Ceci dit, il est possible, et même probable, que les descendants de Primo ancestor soit passé en Afrique par le bassin méditerranéen dès avant 14 millions d'années BP. Ils y ont probablement trouvé, tout comme en Eurasie méridionale d'ailleurs, des descendants du protohominoïde bipède ayant émigré d'Asie dès l'Eocène - proconsuls, morotopithèques, kenyapithèques, dryopithèques, pliopithèques, etc. - et ils se sont imposés au sol par rapport à ces derniers, les poussant à se réfugier dans les arbres, selon le même scénario que dans le berceau original sud-asiatique de l'espèce. Il est probable aussi que, dès cette époque, ces descendants de Primo ancestor avaient commencé à se séparer en trois sous-espèces, une toujours préférentiellement bipède et deux plus arboricoles, ancêtres des gorilles et des chimpanzés actuels, si l'on en juge par les dents morphologiquement proches de celle des chimpanzés, datées de 12,5 millions d'années BP, retrouvées récemment sur le site de Ngorora au Kenya. Néanmoins, des hybridations étaient certainement possibles, et ce jusque la séparation définitive entre la lignée humaine et celle du gorille vers -8 millions d'années, ainsi que jusqu'à -6 millions d'années, pour la séparation entre Homo et Pan.
Quoiqu'il en soit, lorsque vers 8 millions d'années BP, après une impossibilité d'environ 6 millions d'années, les descendants de Primo Ancestor et les sous-espèces apparentées purent à nouveau quitter l'Europe pour coloniser l'Afrique du fait d'un assèchement progressif de la Méditerranée (qui sera presque complètement asséchée vers -5,6 millions d'années avant de se remplir à nouveau, très rapidement, à partir de -5,3 millions d'années du fait de l'ouverture du détroit de Gibraltar), il ne rencontrèrent quasiment que des cercopithèques, des singes à queue arboricoles. L'ensemble des anthropoïdes africains - y compris les représentants de Primo ancestor issus de ses premières incursions sur ce continent avant l'établissement de la barrière marine méditerranéenne - ayant en effet subi une extinction massive du fait des bouleversements climatiques durant cette période. Vers 7 millions d'années BP, ils sont présents dans l'actuel Tchad - comme en témoignent les restes fossiles de Toumaï, probable hybride entre un descendant bipède du Primo ancestor et une sous-espèce arboricole de type "proto-gorille" - et on les retrouve également, vers -6 millions d'années, au Kenya, avec Orrorin tugenensis, descendant de Primo ancestor ayant gagné en taille (1,40m pour 50 kg) et en capacité crânienne par rapport à son ancêtre issu du berceau sud-asiatique. Dans cette même région d'Afrique orientale, on retrouvera d'autres descendants de Primo ancestor, 3 millions d'années plus tard, sous le nom de Homo habilis (ou d'Homo rudolfensis, son proche cousin), ce qui implique que, selon moi, ni Orrorin ni Homo habilis/rudolfensis n'appartiennent en fait au genre Homo ; lequel apparaît à une date encore inconnue, située entre 6 et 2 millions d'années BP dans la Corne de l'Afrique, ainsi qu'on va le voir.
 |
| Orrorin Tugenensis (vue d'artiste) |
En effet, c'est dans cet intervalle de temps que va se produire, sous la conjonction de divers facteurs, la naissance d'une nouvelle espèce amenée à conquérir la planète. Pour résumer l'hypothèse que j'emprunte cette fois-ci (à partir de prémisses différentes) à Elaine Morgan, telle qu'exposée dans son ouvrage Les cicatrices de l'évolution : dans la Corne de l'Afrique, à partir de 16 millions d'années BP, le détachement progressif du bloc Danakil, cette chaîne montagneuse bordée au nord et à l'est par la Mer Rouge et le Golfe d'Aden, ouvre la dépression des Danakil au sud-ouest. Sous l'influence de l'activité volcanique très importante dans la région, cette dépression sera soumise au fil des millions d'année suivantes à des alternances de submersion et d'assèchement. Vers 6 millions d'années BP, à la fin d'une de ces phases d'assèchement, la dépression est à nouveau brutalement remplie par la mer, isolant le bloc Danakil qui deviendra de ce fait une île pendant environ quatre millions d'années, jusqu'au nouvel assèchement suite à la constitution d'un "bouchon" de lave vers 2 millions d'années BP. Isolant, par voie de conséquence, pendant toute cette période, la faune locale "prisonnière" de ce nouveau contexte insulaire, parmi laquelle les descendants de Primo ancestor et les formes apparentées plus ou moins bipèdes ou arboricoles.
 |
| Carte actuelle de la Corne de l'Afrique avec la dépression Danakil entourée en rouge et le bloc Danakil au nord-est |
[Edit du 19.10.2012 : Cette hypothèse de "l'île Danakil", avancée avant Elaine Morgan elle-même, par un auteur nommé Léo La Lumière, est sérieusement remise en cause par certains critiques de la théorie du "singe aquatique", comme par exemple un certain Jim Moore dont le site s'avère très intéressant par la qualité de ses objections à la théorie. Jim Moore - qui ne donne toutefois pas de références précises à ses affirmations - avance que le bloc Danakil n'a jamais été une île mais tout au plus une péninsule restant accrochée au continent africain par des coulées de basalte au sud-est du bloc loin d'être infranchissable par des hominidés (http://www.aquaticape.org/dropped_ideas.html), si cela s'avère juste, peut-être dans ce cas faudrait-il envisager un autre lieu propice, ayant été successivement lui aussi isolé puis relié au continent africain pour l'apparition du genre humain : à ce titre, l'archipel des Dahlak qui appartient aujourd'hui à l'Erythrée pourrait peut-être convenir - hypothèse à vérifier.
Fin de l'Edit].
Suite à la régression de l'environnement boisé sur le bloc Danakil, les espèces principalement arboricoles sont alors progressivement conduites à l'extinction, peut-être "aidées" en cela par le comportement de plus en plus prédateur du descendant bipède de Primo ancestor. En effet, celui-ci, se met probablement à chasser et à se nourrir de ses proches cousins afin d'assurer sa survie dans l'île, entre autres gibiers potentiels. Dans le même temps, il se tourne plus que jamais vers l'environnement marin pour sa subsistance, pêchant poissons, mollusques et coquillages, nouvel apport de protéines et d'éléments nutritifs riches en Oméga3, importants pour le développement cérébral. Durant cette période, ce primate déjà semi-aquatique (ses ancêtres ayant colonisé les forêts tropicales inondées, les marais, les berges de cours d'eau et de lacs, les lagunes et les mangroves), parfait son adaptation en milieu liquide (adaptation qui restera toutefois incomplète), en perdant notamment en partie sa fourrure (voir les autres signes de début d'adaptation aquatique de la lignée humaine dans les ouvrages d'Elaine Morgan). Sa taille augmente également de façon significative. Cependant, s'il n'avait été question que de ces transformations morphologiques, il n"aurait été question que d'une sorte "d'Homo Habilis géant" à petit cerveau, bon nageur (voire plongeur), chasseur, pêcheur, mais potentiellement sans grandes capacités de développement cognitif et culturel. C'est à une mutation majeure, qui va se répandre rapidement par "dérive génétique" dans cette espèce isolée de tout contact avec le continent africain, que l'on doit l'apparition de l'Humanité "véritable", selon moi : le passage de 48 à 46 chromosomes, avec des évolutions morphologiques liées entre autres à des modifications dans les rythmes de croissance par des phénomènes d'hétérochronie et des adaptations à un milieu terrestre très peu boisé. Avec aussi et surtout de nouvelles mutations du gène FOXP2 (mais aussi du gène Srgap2b, voir ici), permettant un "saut quantique" quant au développement du langage, de la cognition, des interactions sociales et des innovations culturelles. De ce fait, le cerveau et la boîte crânienne suivent aussi le mouvement. A noter : c'est pendant cette phase cruciale d'isolement que se répand sur le continent africain, une épidémie virale touchant, à partir des babouins, les autres primates, ce qui explique que le marqueur de cette infection, présent chez tous les grands singes africains, mais pas asiatiques, ne soit pas retrouvé dans le cas de l'espèce humaine (ce qui avait conduit les chercheurs à l'origine de cette découverte à émettre l'hypothèse d'une origine asiatique de l'espèce humaine, comme le signale Elaine Morgan. Voir Benveniste R.E, Todaro G.J., "Evolution of type C viral genes : evidence for an Asian origin of man", Nature, mai 1976, pp. 101–108, cité par Morgan E., Les cicatrices de l'évolution, 10/18, 1990, pp. 232-236).
Aussi, quand l'assèchement progressif de la dépression permet à nouveau le passage depuis le bloc Danakil vers le continent africain, vers 2 millions d'années BP, cette espèce inédite déferle sur les nouveaux territoires qui s'offrent à elle, et, très rapidement, entre en concurrence avec les populations restées en place (avec lesquelles elle ne peut désormais plus se reproduire). A savoir un "buisson" d'espèces hominoïdes, plutôt arboricoles, proches des gorilles : les paranthropes - "buisson" séparé, depuis déjà 6 millions d'années, de la lignée qui donnera Hommes et chimpanzés ; un autre "buisson" d'espèces, plutôt arboricoles elles aussi, proches des chimpanzés : les australopithèques ; et un troisième "buisson" formé par les descendants plutôt bipèdes de Primo ancestor, répartis en deux sous-espèces principales : Homo habilis et Homo rudolfensis, à la face plus ou moins plate. Sans compter - en sachant qu'il n'y a pas une rupture franche entre ces deux derniers "buissons" - de nombreux hybrides, plus ou moins Homo habilis/rudolfensis, plus ou moins australopithèques, avec des caractéristiques en mosaïque (comme c'est le cas, probablement, d'Ardipithécus ramidus, de Kenyanthropus platyops ou d'Australopithécus sediba), liées par ailleurs, pour ce qui concerne l'appareil masticateur, au mode d'alimentation, plus ou moins omnivore, plus ou moins riche en racines coriaces, etc. Toutes ces espèces disparaîtront progressivement, en quelques centaines de milliers d'années, à partir de l'entrée en scène du nouvel arrivant, aux capacités bien supérieures, qui se répandra dans toute l'Afrique, puis sur l'ensemble de la planète. Du côté est de la Rift Valley, ces populations ne donneront de ce fait pas naissance aux espèces de grands singes africains existant encore aujourd'hui (c'est là que fameuse vallée du Rift au coeur de la théorie de la Coppens intervient dans mon hypothèse). Alors qu'à l'ouest de cette vallée, dans les espaces de forêt tropicale préservés, si les hominoïdes bipèdes descendants de Primo ancestor disparaissent, eux aussi, peut-être victimes, après leurs congénères de l'est, de l'arrivée de leurs nouveaux concurrents, les espèces cousines plus arboricoles vont être presque complètement reléguées dans les arbres où elles vont donner nos gorilles et chimpanzés actuels.
A quoi ressemblent ces premiers humains "véritables"? Ils sont morphologiquement beaucoup plus proches de nous que de leurs ancêtres préhumains, atteignant une taille respectable, dans une fourchette comprise - on peut le supposer - entre 1,40m et 1,70m, voire 1,80m pour les mâles adultes. Ils sont certainement - si l'hypothèse "semi-aquatique" est exacte - déjà quasi "nus" et foncés de peau. Leur capacité crânienne varie, mais est très probablement comprise entre 800 et 1300 cm3, peut-être même plus. Mais surtout, ce sont désormais de véritables bipèdes "tout-terrain", aux pieds arqués, avec voûte plantaire, faits pour la marche rapide et pour la course. Ils ont grandement complexifié le langage et les techniques rudimentaires (industrie lithique, notamment) héritées de leurs ancêtres préhumains. Peut-être, même, maîtrisent-il l'utilisation du feu... Un de leurs représentants les plus proches, chronologiquement parlant du début de l'expansion de l'espèce est certainement Homo ergaster (entre 1,50m et 1,80m pour 55kg, et de 800 à 1000 cms3 de capacité crânienne, vivant en Afrique de 1,8 à 1 millions d'années BP) ... Mais il n'est peut-être pas le plus ancien parmi les fossiles que l'on connaît aujourd'hui.
 |
| Crâne d'Homo ergaster |
En effet, selon moi, le fameux crâne dit "de Broken Hill" ou "de Kabwe", associé à l'hominidé que l'on a dénommé Homo rhodesiensis, pourrait bien être beaucoup plus ancien que son estimation actuelle par les paléoanthropologues. Pour l'anecdote, signalons que ce crâne, découvert en 1921 dans un grotte exploitée comme mine de fer et de zinc dans l'ex-Rhodésie, est présenté, à l'inverse comme très récent par des créationnistes et aussi par quelques cryptozoologues, dans la mesure où il est le seul fossile antérieur au néolithique à présenter des caries dentaires (une pathologie censée être apparue avec l'agriculture). De plus, il présente la marque d'un petit trou rond réossifié sur l"écaille temporale gauche que certains considèrent comme le résultat d'une blessure par arme à feu! Pour ma part, j'estime que les explications avancées à ce propos par la science officielle sont plausibles (pour les caries : une intoxication au plomb de l'individu concerné durant son existence qui auraient fragilisé les dents, et pour la lésion temporale, une ostéite).
 |
| Crâne de l'Homme de Kabwe (Homo rhodesiensis) |
A contrario, je suis convaincu que ce crâne date en vérité de 1,75 à 2,5 millions, comme certaines études l'ont, semble-t-il, montré (voir ici, mais je n'ai pas retrouvé les références précises sur lesquelles se fonde cette information). Les paléoanthropologues refusent pourtant cette estimation et ne lui assignent aujourd'hui, généralement, que quelques centaines de milliers d'années. Toutefois, cette datation n'a pas été établie de façon absolue, ni même par rapport à l'environnement dans lequel ce fossile a été trouvé (ce qui aurait été impossible du fait des conditions de sa découverte), mais relativement à la capacité crânienne de presque 1300 cms3 paraissant beaucoup plus en adéquation avec un pré-sapiens. Pour moi, il s'agit en fait d'un des tout premiers représentants du genre Homo, peu de temps après leur exode hors du bloc Danakil, portant des caractéristiques aussi bien d'Homo erectus que de Néandertal ou de Sapiens, puisque bien antérieur à la séparation de ces trois lignées humaines. Ce crâne est assez similaire à d'autres, aussi peu spécialisés, dont l'âge n'est pas connu avec certitude, comme ceux de Bodo, en Ethiopie (- 500 000 ans?), ou de Petralona en Grèce (- 700 000 ans?).
En résumé, d'après ma théorie, le genre humain à proprement parler, avec ses 46 chromosomes, sa capacité à la marche rapide et à la course et ses compétences cognitives et langagière, est apparu voilà environ 2 millions d'années dans la Corne de l'Afrique après s'être développé de façon totalement isolée pendant 3 ou 4 millions d'années dans l'île formée alors par le bloc Danakil. En expansion à partir de ce point focal, Homo a éliminé en quelques centaines de milliers d'années tous ses concurrents locaux plus ou moins bipèdes, issus d'ancêtres communs, rencontrés en chemin. Très rapidement, il s'est répandu en Asie et en Europe où il a été sans doute là aussi responsable, directement ou indirectement, de l'extinction d'un certain nombre d'espèces rencontrées, descendantes de Primo ancestor, comme par exemple Homo georgicus dont on a découvert, dans l'actuelle Georgie, des restes datés de 1,8 millions d'années et qui est pour moi, une sorte d'"Homo habilis européen", un descendant de Primo ancestor un peu évolué, présent là depuis peut-être 10 ou 15 millions d'années, bien avant l'arrivée,en tout cas, des véritables représentants de la lignée humaine. On rencontre la même situation, très probablement, dans le cas du prétendu "Homme de Wushan" en Asie, dont on a trouvé une mandibule datée de 1,8 millions d'années BP sur le site de Longuppo en Chine. C'est certainement à cet autre descendant, asiatique celui-là, de primo ancestor, que peut être associée une industrie lithique, trouvée sur le site voisin de Renzidong, dont l'ancienneté est estimée à 2,2 millions d'années BP. Dans certains cas, cependant, il s'agit plutôt d'une "quasi" extinction, puisque la présence d'Homo floresiensis au paléolithique, voire dans les temps historiques en Asie du sud-est, témoigne de la persistance de petits groupes résiduels issus de certaines de ces espèces plus ou moins anciennes - très ancienne, là, en l'occurrence - dans des "poches" géographiques relativement isolées. Il est plus que probable, d'ailleurs, qu'une population résiduelle étroitement apparentée à celle de Florès (la souche-mère d'Homo Floresiensis selon moi) survive encore aujourd'hui à Sumatra où elle est connue sous le nom d'Orang pendek ou de sedapa (en supposant qu'un petit groupe soit arrivé à Florès vers 1,2 millions d'années BP sur des radeaux flottants). [Edit du 30.06.2012 : Des créatures très similaires ont été décrites par de nombreuses populations dans toute l'Asie du sud-est sous divers noms (batutut, segoegoe, nguoi rung, etc.), jusqu'au Sri Lanka. Particulièrement intéressant pour notre propos : elle n'est pas inconnue non plus dans l'archipel des Mentawaï, qui constitue selon moi le berceau de l'espèce, et partant de là, le berceau de toute la lignée des primates hominoïdes (voir ci-dessus). Fin de l'Edit].
 |
| Orang pendek de Sumatra (vue d'artiste) |
"Le 'Hobbit' nous dit-il que le véritable berceau de l'humanité est l'Asie?"
Ce commentaire, extrait de cet excellent reportage sur l'Homme de Florès (44:26), ouvre une dernière partie
passionnante par ses questionnements où il est question, entre autres, d'Homo georgicus
Sortis très tôt du continent africain, vers 1,9 millions d'années BP, les premiers représentants du genre Homo se sont progressivement spécialisés en Asie pour donner la lignée des Homo erectus (1,6 millions d'années BP pour les premiers restes fossiles appartenant sans ambiguïté au genre Homo, trouvés sur le site indonésien de Sangiran - je reste plus dubitatif pour la calotte cranienne de l'"enfant de Mojokerto" qui pourrait bien avoir appartenu plutôt à un descendant de Primo ancestor, tout comme pour la mandibule trouvée sur le site chinois de Longgupo) et, en Europe, celle des Homo antecessor (présence attestée en Espace sur le site d'Atapuerca vers 1,2 millions d'années BP). Une deuxième vague, vers -700 000 ans donnera les Homo heidelbergensis en Europe. Puis, une troisième migration, vers -400 000/-300 000 ans verra l'arrivée de l'Homme de Denisova en Asie et de son proche cousin Néandertal en Europe, qui se métisseront avec leurs prédécesseurs cités ci-dessus (d'où, par exemple, les incisives "en forme de pelle" que l'on retrouve en Asie, chez les Homo erectus comme chez les sapiens asiatiques actuels, caractère très probablement transmis par le biais des Denisoviens dont les toutes premières populations arrivées en Asie se sont sans doute métissées avec des erectus, les descendants en question se métissant eux, bien des générations plus tard, avec des sapiens). La quatrième et dernière vague sera celle, justement, des Homo sapiens qui, suite à la quasi éradication de l'espèce Homo de la planète, vers 74 000 ans BP, du fait de changements climatiques drastiques dus aux conséquences de l'explosion du supervolcan asiatique Toba sur l'île de Sumatra, se répandront dans le monde entier à partir d'une petite population résiduelle totalement "sapientisée" en Afrique australe, submergeant en chemin les autres populations survivantes, très réduites numériquement, tout en se mélangeant avec elles, se métissant même, a minima, avec celles déjà assez voire très spécialisées sur d'autres continents (avec les Néandertaliens en Europe et au Proche-Orient et, comme dit plus haut, avec les Dénisoviens en Asie).
Ce métissage eut peut-être même lieu, très occasionnellement, avec les Néandertaliens les plus occidentaux dans l'espace européen, malgré l'hyperspécialisation de ces derniers, associée sans doute, déjà à l'époque de la rencontre, à un phénomène de régression évolutive, cause, finalement, de leur (quasi?) disparition, comme certains indices le laissent supposer. Si un article (voir ici pour la version française), publié originellement en espagnol, en 2006, par un zoologue, Juan-Luis Domenech Quesada dans la revue cryptozoologique Bipedia éditée par François de Sarre soutient une telle thèse, c'est toutefois la science "officielle" elle-même qui a mis en évidence, de façon formelle, le principal indicateur de cette régression : non seulement la croissance des Néandertaliens (surtout, probablement, ceux de la période tardive) s'avère avoir été beaucoup plus rapide que celle des Sapiens, mais, étonnamment, elle était aussi plus rapide que celle de leurs prédécesseurs, Homo heidelbergensis ou même Homo antecessor, comme l'ont montré diverses études effectuées à partir de 2004 sur les dents des espèces en question (voir ici ou là), se rapprochant de celle du chimpanzé (Néandertal était adulte à 15 ans, contre 12 ans pour le chimpanzé et 18 pour l'Homo sapiens). Cette différence irréductible avec Homo sapiens, sans doute consécutive à un stress environnemental et à la réduction drastique des effectifs entraînés par la glaciation consécutive à l'explosion du supervolcan Toba, semblait, à l'époque des premières études, en 2004, appuyer l'hypothèse de la non-hybridation Néandertal/Sapiens, a priori impossible du fait d'une trop grande divergence spécifique. Pourtant, on sait aujourd'hui, grâce au décryptage des génomes néandertaliens et sapiens que tous les Humains actuels, sauf africains, portent entre 1% et 4% de gènes légués par notre cousin européen. Comment expliquer cette apparente contradiction? Je propose une réponse à cette question dans le récapitulatif suivant qui constitue :
Un tableau général des diverses vagues de migrations pour tenter de mieux comprendre ce qui a dû se produire depuis 2 millions d'années :
-2 millions d'années : Homo (non spécialisé) se répand à partir de la corne de l'Afrique dans toutes les directions, notamment vers le sud et vers le nord. Il possède encore une grande diversité génétique et une grande variabilité morphotypique que ce soit pour la taille corporelle, la capacité cérébrale ou les traits généraux qui peuvent soit évoquer des caractères qui seront définis plus tard de façon subspécifique puis spécifique comme "néandertaliens", "erectus" (exemple d'Homo ergaster que certains spécialistes voient comme un Homo erectus africain) ou "sapiens", soit rassembler ces trois types selon des configurations variables (comme chez l'homme de Kabwe).
-1,9 millions d'années : cet Homo non spécialisé sort d'Afrique pour se répandre en Asie et en Europe (évènement que l'on nomme habituellement "Out of Africa 1"). Il y retrouve les descendants non humains de son ancêtre Primo ancestor, en élimine, directement ou indirectement, une grande partie et pousse les survivants à se réfugier dans des espaces reculés (montagnes, forêts), voire pour certains à abandonner presque complètement la bipédie pour l'arboricolisme (cas des orang-outans, similaire à celui des gorilles et des chimpanzés en Afrique?)
- 1,2 millions d'années : la spéciation (et spécialisation) par distance est enclenchée aux marges les plus éloignées du point focal constituée par la corne de l'Afrique (Europe occidentale, Chine et Asie du sud-est ) : "néandertalisation" en Europe de l'ouest, "erectisation" en Asie de l'est, "sapientisation" en Afrique du sud. Ces phénomènes vont toucher de façon continue au fil des centaines de millénaires les vagues successives de migration qui se métisseront les unes avec les autres (en Europe, la première vague, celle d'Homo antecessor se métissera avec la vague suivante pour donner Homo heidelbergensis, puis les Homo heidelbergensis se métisseront avec les Homo de la vague suivante pour donner Homo néandertalensis ; en Asie, les représentants de la première vague de migration ayant évolué vers la forme Homo erectus, ceux-ci se métisseront avec les Hommes de la seconde vague pour donner les Dénisoviens). Quant à H. cepranensis, hominidé dont un reste fossile fut découvert en Italie en 1994 et daté de 900 000 ans, il est peut-être une variété régionale d'Homo antecessor, ou résulte peut-être du métissage entre l'Homo erectus asiatique (certains groupes ayant pu migrer en Europe) et Homo antecessor...
-75 000 ans : Ce phénomène de spéciation par distance est maintenant bien en place, néanmoins des échanges de flux géniques sont encore possibles de proche en proche (selon le phénomène dit "du gradient", voir ici, l'exposition de l'hypothèse en question appliquée aux Homo néandertalensis par le paléoanthropologue Jean-Luc Voisin). Cela signifie toutefois que les populations les plus en marge de l'immense espace occupé désormais par le genre Homo, de l'extrême sud du continent africain au confins de l'Europe occidentale et de l'Asie orientale n'échangent quasiment plus de gènes : celles d'Afrique australe sont désormais "sapiens", celles d'Europe occidentale sont constituées de Néandertaliens aux caractères physiques bien affirmées, et enfin, celles d'Asie de l'est sont des Dénisoviens plus ou moins nettement "erectisés" (cas des ancêtres locaux des hominidés de la grotte du Cerf rouge en Chine ou de l'Homme de Solo sur l'île de Java?), même s'il se rencontre encore également des individus avec certaines caractéristiques autres, néandertaliennes ou sapiens comme en témoignent un nombre grandissant de fossiles chinois tels que le crâne de Dali.dont l'ancienneté est estimée à environ 230 000 ans et surtout les restes des hommes de Zhirendong (100 000 ans BP), de Xuchang (80 000ans BP) ou de Liujiang ( au moins 68 000ans), tous trois de type nettement sapiens.
- 74 000 ans : l'explosion du supervolcan Toba sur l'île de Sumatra induit d'énormes répercussions sur les écosystèmes d'une bande territoriale allant de l'Asie du sud à l'Europe méridionale, incluant également tout le nord du continent africain, en provoquant une baisse des températures moyenne de plusieurs degrés. Cette période de glaciation qui dure, semble-t-il, environ 20 000 ans provoque une diminution drastique des diverses populations d'Homo et de pré-Homo dans ces territoires, mais non leur disparition complète - y compris à Sumatra même et dans les îles voisines, pourtant très proches du lieu de l'éruption puisque, au moins pour ce qui concerne les descendants directs de Primo ancestor (Homo floresiensis), ces derniers y ont survécu encore plusieurs dizaines de milliers d'années (c'est le cas peut-être même encore aujourd'hui à Sumatra sous la forme d'Orang pendek). Les populations partiellement sapiens survivantes du Proche et du Moyen-Orient refluent vers le sud, vers l'Afrique, remplacées localement par des Néandertaliens de type plus ou moins "marqué" (comme ceux de la grotte de Kebara en Israël, dont les restes fossiles ont été datés entre 60 000 et 48 000 ans BP) venus des territoires plus septentrionaux (d'Europe méridionale, ceux d'Europe occidentale et septentrionale resteront "coincés" dans leur cul-de-sac géographique pour finalement - quasiment? - disparaître).
 |
| Intérieur de la grotte de Blombos (Afrique du sud) |
Une population, quant à elle, est complètement préservée, tout au sud de l'Afrique. Il s'agit d'hommes anatomiquement modernes, des Homo sapiens très probablement noirs de peau, en avance culturellement et technologiquement à l'époque sur le reste de l'Humanité, comme en témoignent les étonnants vestiges datés de 80 000 à 70 000 ans BP dans la grotte de Blombos (située actuellement près de la ville du Cap). Outre des outils en pierre obtenus par des techniques qui ne seront connues que 50 000 ans plus tard en Europe, on a trouvé aussi des parures de coquillages, et des matériaux utilisés par des activités décoratives, dont des blocs d'ocre gravés, premiers signes de ce genre d'activité de la part de l'humanité. Ils furent sans doute aussi les premiers humains à se vêtir (l'apparition du vêtement aurait eu lieu voilà environ 72 000 ans en Afrique selon une étude réalisée sur les... poux liés au genre humain. voir ici, ce qui leur aurait permis de coloniser les régions septentrionales du globe - pour un complément sur les histoires de poux liées aux hominidés, voir aussi ici) ainsi que les premiers à réaliser des peintures rupestres, comme celles trouvées par milliers en Namibie, par exemple, même si les plus anciens exemples connues - pour l'instant - en la matière ne remontent "qu'à" 26 000 ans BP (grotte Apollo 11 en Namibie). Leurs descendants actuels sont probablement les "San", aussi connus sous le nom de "Bushmen", dont la civilisation semble bien éloignée de celle de leurs lointain ancêtres, lesquels, après une apogée probable voilà plusieurs dizaines de milliers d'années, connurent certainement un déclin progressif, accentué par les coups de boutoir des colons européens à partir du XVIIIe siècle.
 |
| Lames de silcrète (grotte de Blombos, Afrique du sud) |
 |
| Bloc d'ocre gravée (grotte de Blombos, Afrique du sud) |
-70 000 ans : Expansion vers le nord, par voie terrestre ainsi que par navigation le long des côtes, de ces Homo sapiens d'Afrique australe. En 10 000 à 15 000 ans à peine ils colonisent ainsi une grande partie du continent et s'installent, vers - 50 000 BP au Proche-orient (Out of Africa 2, voir ici la confirmation de cette hypothèse par des études génétiques récentes datées de 2009), où ils côtoient certainement des Néandertaliens assez peu spécialisés avec lesquels les métissages sont encore occasionnellement possibles, avant de continuer leur progression vers l'Europe d'une part, vers l'Asie d'autre part, en perdant progressivement, mais assez rapidement, du fait des métissages et des mutations, leurs caractères "négroïdes". Arrivés à l'ouest de l'Europe vers 40 000 ans BP, ils vont y déployer tout leur génie créatif et artistique sur les parois de grottes comme Chauvet (-33 000 ans) ou Lascaux et dans bien d'autres manifestations culturelles comme les Vénus stéatopyges (qui rappellent le morphotype sud-africain d'origine). Ils cohabitent là, pendant plusieurs milliers d'années avec des Néandertaliens très spécialisés, lesquels sont en rapide régression selon un processus - proposé en son temps par le zoologue Bernard Heuvelmans - de "déshumanisation" (régression marquée par le raccourcissement important du temps de croissance évoqué plus haut, mais aussi régression culturelle). Les métissages entre les Homo sapiens et ces Néandertaliens d'Europe occidentale ne sont alors quasiment plus possibles, ou alors peut-être pour donner, le plus souvent, des "hybrides" eux-mêmes inféconds. A l'est, le métissage avec les Dénisoviens semble avoir été encore plus rare, mais les Hommes dits "de la grotte du Cerf Rouge" pourraient être issus de ces mélanges (voir ici). Une branche de cette population métissée pourrait par ailleurs avoir contribué au peuplement de l'Australie et de la Papouasie Nouvelle-Guinée (voir ci-après) puisque l'on a retrouvé des gènes dénisoviens dans les populations humaines actuelles de cette région du monde.
-65 000 ans : Les Sapiens d'Afrique du Sud ayant été, semble-t-il, jusqu'à inventer - même si cela peut paraître difficile à croire - l'embarcation à proue haute permettant la navigation en haute mer, on les retrouve loin de l'espace géographique d'origine, à l'est et à l'ouest, dans des endroits inattendus. S'ils n'ont apparemment pas accosté à Madagascar (les courants océaniques ne les ayant peut-être pas mis sur la route maritime de la Grande Île), ils abordent, beaucoup plus loin, sur les rivages australiens. Ils vont s'y installer pour constituer le premier peuplement de l'île-continent. Il s'agit des Hommes dits "de Mungo" (restes fossiles datés de 40 000 ans BP - voire 60 000 ans - trouvés au bord du lac asséché de Mungo en Nouvelle Galles du sud) auxquels sont associées des peintures rupestres de la Terre d'Arnhem dont la plus ancienne estimée est celle d'un émeu disparu localement depuis 45 000 ans (Gunn, Douglas, Whear, 2011). A noter qu'une des plus anciennes peintures rupestres locales (à Kimberley), datée d'au moins 17 000 ans BP, représente une pirogue géante à proue haute, embarcation inconnue des aborigènes découverts par les occidentaux au XVIIIe siècle...
 |
| Une des peintures rupestres dites "de Bradshaw" (Kimberley, Australie). Age estimé : au moins 17 000 ans |
[Edit du 05.07.2012 : Voir ici un site sur les étonnantes peintures rupestres dites "Bradshaw" (du nom du découvreur) ou "Gwion Gwion" (selon le nom donné par les aborigènes actuels) de Kimberley, dont certaines pourraient dater de plus de 46 000 ans... Fin de l'Edit]
Ces Hommes de Mungo, d'aspect "gracile", furent rejoints quelques 20 000 ou 30 000 ans plus tard par des Homo sapiens de type "australoïde", descendants de migrants africains sur le continent asiatique qui, du fait de métissages probables avec diverses populations lors de leur périple (avec des H. sapiens et des Néandertaliens généralistes, mais aussi avec des Dénisoviens en voie d'extinction), avaient acquis un aspect beaucoup beaucoup robuste. Des restes fossiles d'individus issus de cette nouvelle vague d'immigration en Australie ont été trouvé notamment sur le site de Kow Swamp.
Il est par ailleurs possible que des hommes identiques à ceux de Mungo se soient installés, suite là aussi à des périples maritimes, plus haut que l'Australie dans divers endroits en Asie du sud-est : leurs descendants étant, selon moi, les peuples dits "Négritos" qui vivent encore aujourd'hui dans les îles Andaman, ainsi que dans certaines régions des Philippines et de la péninsule malaise. Même si ces peuples ont préservé grosso modo le morphotype originel, leur génome a évolué pendant plusieurs dizaines de millénaires de façon totalement isolée par rapport à la souche d'origine, ce qui explique qu'ils sont loin, génétiquement, des sud-africains actuels, d'autant qu'ils ont subi l'influence de métissages, probablement assez récents, avec des populations austronésiennes.
A l'Ouest, c'est en Amérique du sud que l'on retrouve des sites très anciens, datant de cette période (notamment le plus ancien aujourd'hui découvert : celui de Pedra Furada au Brésil, daté de -60 000/-55 000 ans, voir ici, même si ces datations sont encore controversées où l'on a trouvé là aussi, comme à Kimberley une peinture rupestre représentant un bateau. De nombreux restes humains de type "négroïdes" datés entre 35 000 ans et 11 000 ans BP, ont également été trouvés au Brésil comme, sur le site de la grotte de Lapa Vermelha, celui d'une femme d'environ 25 ans qui fut dénommée Luzia, preuve que le premier peuplement de l'Amérique (du sud en l'occurrence) fut le fait d'une population en provenance du continent africain (quoique certains voient pour les restes fossiles comme ceux de Luzia une origine australoïde, par le biais de groupes en provenance d'Océanie). Des traces de présence humaine (restes fossiles, foyers, outils, empreintes, etc) aussi voire plus anciennement datés ont été également trouvés récemment au Chili, au Mexique ou au Pérou (voir l'article très complet de wikipédia pour toutes ces informations. Voir également ici (autre article wikipédia) pour des informations plus générales sur le premier peuplement des Amériques.
 |
| Reconstitution du visage de Luzia par tomographie informatique |
 |
| Luzia, modelé par l'artiste Cicero Moraes (image sur l'article wikipédia) |
-20 000 ans : Les caractéristiques physiques générales (africaines, caucasiennes, mongoloïdes, australoïdes) sont désormais en place sur les divers continents, la situation se complexifiant localement du fait de multiples migrations multipolaires et multidirectionnelles régionales avec les métissages subséquents. Une population asiatique mongoloïdes arrive en Amérique du Nord par le détroit de Behring : souche ancestrale des Amérindiens actuels, mais d'autres populations (caucasiennes - voir l'Homme de Kennewick - et peut-autre aussi australoïdes ainsi que, plus tardivement, austronésiennes) semblent avoir également contribué au peuplement des Amériques.
-11 000 ans : Arrivée de populations austronésiennes modernes sur l'île de Florès en Indonésie.
1923 : Rencontre d'un Orang pendek en forêt de Sumatra par un Hollandais, Van Herwaarden (qu'il décrit, de façon détaillée, sous le nom de "sedapa"), rapporté en 1961 par le cryptozoologue Ivan Sanderson (2008, pp. 222-223) :
"Then I happened by chance to look round to the left and spotted a slight movement in a small tree that stood alone. By now it was time for me to be going home, for it was not advisable to journey through such country after sundown. But all the same I was tempted out of curiosity to go and see what had caused the movement I had noticed. What sort of animal could be in that tree? My first quick look revealed nothing. But after walking round the tree again, I discovered a dark and hairy creature on a branch, the front of its body pressed tightly against the tree. It looked as if it were trying to make itself inconspicuous and felt that it was about to be discovered.
It must be a sedapa. Hunters will understand the excitement that possessed me. At first I merely watched and examined the beast which still clung motionless to the tree. While I kept my gun ready to fire, I tried to attract the sedapa's attention, by calling to it, but it would not budge. What was I to do? I could not get help to capture the beast. And as time was running short I was obliged to tackle it myself. I tried kicking the trunk of the tree, without the least result. I laid my gun on the ground and tried to get nearer the animal. I had hardly climbed 3 or 4 feet into the tree when the body above me began to move. The creature lifted itself a little from the branch and leant over the side so that I could then see its hair, its forehead and a pair of eyes which stared at me. Its movements had at first been slow and cautious, but as soon as thesedapa saw me the whole situation changed. It became nervous and trembled all over its body. In order to see it better I slid down on to the ground again.
The sedapa was also hairy on the front of its body; the colour there was a little lighter than on the back. The very dark hair on its head fell to just below the shoulder-blades or even almost to the waist. It was fairly thick and very shaggy. The lower part of its face seemed to end in more of a point than a man's; this brown face was almost hairless, whilst its forehead seemed to be high rather than low. Its eyebrows were the same colour as its hair and were very bushy. The eyes were frankly moving; they were of the darkest colour, very lively, and like human eyes. The nose was broad with fairly large nostrils, but in no way clumsy; it reminded me a little of a Kaffir's. Its lips were quite ordinary, but the width of its mouth was strikingly wide when open. Its canines showed clearly from time to time as its mouth twitched nervously. They seemed fairly large to me, at all events they were more developed than a man's. The incisors were regular. The colour of the teeth was yellowish white. Its chin was somewhat receding. For a moment, during a quick movement, I was able to see its right ear which was exactly like a little human ear. Its hands were slightly hairy on the back. Had it been standing, its arms would have reached to a little above its knees; they were therefore long, but its legs seemed to me rather short. I did not see its feet, but I did see some toes which were shaped in a very normal manner. This specimen was of the female sex and about 5 feet high.
There was nothing repulsive or ugly about its face, nor was it at all ape-like, although the quick nervous movements of its eyes and mouth were very like those of a monkey in distress. I began to walk in a calm and friendly way to the sedapa, as if I were soothing a frightened dog or horse; but it did not make much difference. When I raised my gun to the little female I heard a plaintive "hu-hu," which was at once answered by similar echoes in the forest nearby.
I laid down my gun and climbed into the tree again. I had almost reached the foot of the bough when the sedapa ran very fast out along the branch, which bent heavily, hung on to the end and then dropped a good 10 feet to the ground. I slid hastily back to the ground, but before I could reach my gun again, the beast was almost 30 yards away. It went on running and gave a sort of whistle. Many people may think me childish if I say that when I saw its flying hair in the sights I did not pull the trigger. I suddenly felt that I was going to commit murder. I lifted my gun to my shoulder again, but once more my courage failed me. As far as I could see, its feet were broad and short, but that the sedapa runs with its heels foremost is quite untrue."
Années 1990 : Premières enquêtes de terrain approfondies sur l'Orang pendek par les journalistes britanniques Deborah Martyr et Jeremy Holden.
2003 : Découverte de restes d'individus hominoïdes inconnus de très petite taille, surnommés plus tard "Hobbits"(autrement dit "Homo floresiensis), dans la grotte Liang Bua sur l'île de Flores...
 |
| Grotte de Liang Bua, Florès, Indonésie |
[Edit du 07.10.2010 :
Trouvé sur le site www.ma.prehistoire.free.fr, ce compte-rendu, par l'auteur du site, d'un certain nombre de remarques "de bon sens" de l'éminent paléoanthropologue John Hawks, dont j'ai déjà mis en lien depuis plusieurs mois le blog passionnant (en anglais). Ces remarques me semblent tout à fait conforter mon hypothèse au sujet de l'"Homme" de Florès :
John Hawks (Université du Wisconsin) a émis quelques remarques :
- LB1 [le premier spécimen touvé dans la grotte de Liang Bua] a la taille et le volume cérébral d'un Australopithèque, mais H. floresiensis est trop éloigné géographiquement et chronologiquement du dernier Australopithèque pour être attribué à ce genre ;
- l'île de Flores n'a jamais été reliée au continent : faut-il conclure à l'existence de la navigation il y a 800 000 ans ?
- le seul argument pour faire de l'H. erectus javanais l'ancêtre de LB1 est sa proximité géographique et son antériorité dans le temps ;
- LB1 n'est pas un erectus : il ne possède en effet qu'un seul des huit caractères qui définissent erectus (la largeur maximale du crâne est située au niveau de a base du crâne)
- quand un Mammifère évolue vers un individu de taille réduite, le volume de son cerveau ne diminue pas d'autant (pour une réduction de taille de 50%, le cerveau ne perd que 10% de son volume) : le type de nanisme de LB1 avec cette microcéphalie est inhabituel, surtout pour un Primate.
- LB1 est une femme ; la taille moyenne de sa population est sans doute plus grande ;
- LB1 est-il un individu pathologique ? Habituellement, les personnes de petite taille ont un cerveau de volume normal ; les individus microcéphales (cerveau anormalement réduit) ont habituellement une taille normale ; il ne peut pas être exclu que LB1 soit un individu pathologique, mais aucun argument ne permet actuellement de conclure.
- Hawks propose finalement que LB1 soit considéré comme l'holotype d'une espèce Hobbit floresiensis appartenant à un genre nouveau représenté également par Homo habilis qu'il rebaptise Hobbit habilis pour en faire l'ancêtre de LB1.
Fin de l'Edit]
[Edit du 08/10/2012. Ai trouvé aujourd'hui une page d'un paléoanthropologue réputé, Dennis Etler (http://sinanthropus.blogspot.fr/2010/07/will-real-h-floresiensis-please-stand.html), qui se laisse aller à quelques "rêveries" à propos d'Homo floresiensis, où l'on retrouve l'orang pendek, l'ebu gogo, mais aussi un primate asiatique du Miocène, le lufengpithèque, sans compter des orang-outans géants bipèdes... En résumé, ce chercheur avance l'hypothèse "un peu folle" que H. florensiensis ne serait peut-être pas un Homo du tout, mais une espèce ayant évolué en Asie de façon autochtone à partir du lufengpithèque (il compare des restes fossiles de lufengpithecus à ceux de Liang Bua), suggérant de fait de le renommer "microanthropus floresiensis"... Ce n'est pas tout à fait mon hypothèse, mais ça en est proche, étonnamment proche... Je me sens moins seul, d'un coup! Voir aussi cet autre article sur son blog passionnant, où il s'interroge sur ce lufengpithecus qui pourrait avoir préfiguré, dès le Miocène, certaines caractéristiques attribuées à la lignée humaine : une certaine forme de bipédie, au moins aussi bien que Toumaï ou Orrorin en Afrique, et même, dit-il, la fabrication et l'utilisation d'un outillage lithique : http://sinanthropus.blogspot.fr/2010/07/could-there-be-non-hominan-hominids.html. Il en tire l'hypothèse que lufengpithecus aurait pu être à l'origine d'une évolution au sein des pongidés, parallèle à celle qui eut lieu à la même époque, à partir de 7 ou 8 millions d'années BP, en Afrique au sein des hominidés, vers la bipédie et d'autres caractéristiques associées habituellement exclusivement à la lignée humaine (d'où son autre post, cité plus haut sur H. floresiensis comme éventuel descendant bipède du lufengpithèque). Personnellement, mon hypothèse sur une origine encore beaucoup plus précoce de la bipédie, qui serait commune à la lignée ayant abouti aux créatures - non-humaines - de Florès et à la lignée humaine) me satisfait mieux en me semblant plus conforme au principe de parcimonie...
Dans le même ordre d'idées, j'ai trouvé aussi aujourd'hui cet article extrêmement intéressant, paru dans la revue Nature (http://www.nature.com/nature/journal/v459/n7249/full/459910a.html), où un autre chercheur (Russell Ciochon) avoue abandonner la thèse qu'il avait longtemps défendue à propos de la mandibule de Longgupo en Chine en attribuant ce reste fossile à un Homo erectus - ce qui impliquait une présence très ancienne, aux alentours de 2 millions d'années BP - de la lignée humaine en Asie. Il attribue aujourd'hui ce vestige à un "singe mystère" qui ne serait pas un ancêtre d'Orang-outan... mais peut-être un lufengpithèque ou un de ses descendants (voir ici, sur l'excellent site du chercheur John Hawks)... En sachant qu'on a retrouvé un outillage lithique datant apparemment de la même période sur le site, outillage qui ne serait d'ailleurs pas le plus ancien retrouvé en Asie puisque des outils retrouvés sur un autre site chinois, à Renzidong, ont été datés de 2,2 à 2,6 millions d'années... Lufengpithecus serait-il Primo ancestor ou, en tout cas, un descendant proche de celui-ci comme le suggère mon hypothèse? Même si tout cela ne constitue pas des preuves, voilà un beau faisceau de présomptions allant dans ce sens.
Fin de l'Edit]
A suivre...
___________________________________________________
Reférences bibliographiques
Benveniste R.E, Todaro G.J., "Evolution of type C viral genes : evidence for an Asian origin of man", Nature, mai 1976, n°13.
Brandon-Jones, D., "Pre-glacial Bornean primate impoverishment and Wallace’s line", in: Hall, R., Holloway, J.D. (Eds.), Biogeography and Geological Evolution of Southeast Asia, Backhuys Publishers, Leiden, The Netherlands, 1998, pp. 393–404.
Ciochon R., "The mystery ape of Pleistocene Asia", Nature, 459, pp. 910-911, juin 2009.
Deloison Y., Préhistoire du piéton. Essai sur les nouvelles origines de l'homme, Paris, Plon, 2004.
Domenech Quesada J.L., "El 'origen remoto' del Homo sapiens : Una theoria alternativa de la evolucion humana", Bipedia n°24, janvier 2006.
Gunn R.G., Douglas L.C., Whear R.L., "What Bird is that ?" Australian archaeology, n°73, 2011.
Morgan E., Les cicatrices de l'évolution, Paris, 10/18, 1990.
Kittler R,, Kayser M,, Stoneking M., "Molecular evolution of Pediculus humanus and the origin of clothing", Curr Biol, 2003, 13, pp. 1414-1417.
Ramirez Rossi F.V., Bermudez de Castro J.M., "Surprisingly rapid growth in Neanderthals", Nature 428, pp. 936-939.
Sanderson I., Abominable Snowmen. Legend Come to Life, Cosimo, 2008 (1961).
Stoll C., Homogenesis. Histoire de l'histoire de l'Homme, Lausanne, Presses polytechniques et universitaires romandes, 2011.
Thackeray J.F., "Comportement animal, magie cynégétique et art rupestre de l'Afrique australe", Afrique et histoire, 2, 2006, pp. 149-160.
Aucun commentaire:
Enregistrer un commentaire